Introduction
Spinal cord injury (SCI) can result in permanent neurologic deficits and devastating consequences [1]. Inflammation and glial scar formation can result in the failure of axon regeneration in addition to SCI [2,3]. The secondary cascade involves inflammation, matrix degradation, neurotoxicity, and glial scarring formation [4,5]. Glial scars are able to express the chondroitin sulfate proteoglycan. In addition to a nonspecific barrier, glial scars consist of active astrocytes. This means that glial scars can stop the growth of injured axons in the lesion chemically [2]. Matrix metalloproteinase 9 (MMP9) is a contributor to glial scar formation [2,6], and myelin protein degradation and macrophage infiltration are the beginning of the breakdown of the blood-spinal cord barrier (BSCB) [3]. The administration of secretomes in the brain could induce the macrophages to secrete interleukin-10 (IL-10) and transforming growth factor-β (TGF-β) and inhibit the synthesis of matrix metalloproteinase [7]. The conventional treatment is conducted using an operative approach with laminectomy, stabilization, and fusion [8]. Recently, researchers have mainly focused on therapies targeted at secondary cascade SCI [4,5,9]. Although the efficacy of stem cells as a neuroprotective agent is very promising, growing evidence has proven the efficacy of secretomes, which is supported more by the paracrine effects of the secretomes rather than the cells themselves [10,11], and after 14 days of stem cell injection in the tail vein of mice, only about 4% of the stem cells were present in the brain with 10% surviving [7]. Moreover, stem cell secretomes can reduce the negative impact of stem cells, including the possibility of malignancy [10]. The effect of the treatment with human neural stem cell (HNSC) secretomes in SCI is unclear. Accordingly, this study aims to investigate the effects of HNSC secretomes on subacute SCI post-laminectomy by analyzing IL-10, MMP9, TGF-β, and Basso–Beattie–Bresnahan (BBB) score locomotors as expressions of neurological recovery. For this purpose, we used a well-established Rattus norvegicus model of SCI contusion-compression [12,13].
Materials and Methods
1. Ethics statement
The study’s protocol was reviewed and approved by the university (REC.1462/UN25.8/KEPK/DL/2021). All rats were approved by the Animal Health Office (no., 503/A.1/0005.B/35.09.325/2020).
2. Study design
This research has a true experimental design. The sample size was counted using the Lemeshow formula (n=5 animals) with correction factors of 20%. The animals were randomly categorized into three groups: (1) normal, (2) SCI+non-secretome, and (3) SCI+secretome. The SCI+secretome group was given 30 μL HNSC secretome intrathecal injection (Th10) 3 days post-SCI/post-laminectomy [14]. All groups had five replications. We conducted observations with a duration of over 35 days.
3. Preparation of human neural stem cell secretomes
The material of the HNSC secretomes was obtained through the installation of stem cells and tissue bank (REC.0059/KEPK/IX/2020). It produced 20 mL HNSC secretomes on June 21, 2021.
HNSC secretome is characterized by neural cells and nestin markers. The neural stem cells were counted as 2×5×106 and derived from adipose mesenchymal stem cell (AD-MSC). The production process of HNSC secretome includes four phases: the thawing and culturing of AD-MSC, differentiation of AD-MSC into the human neural cells in the Gibco media with CD45 and nestin markers, production of human neurogenic metabolites, and dialysis, validation, and packaging of human neurogenic metabolites.
4. Rats and spinal cord injury models
The adult male R. norvegicus pure strain Sprague–Dawley rats (Research Laboratory and Integrated Testing, Gadjah Mada University, Yogyakarta, Indonesia) aged 3–4 months and weighed 300–350 g. Acclimatization was carried out for 7 days by one laborant and two veterinarians at the Animal and Biomedical Laboratory of the Dentistry Faculty, University of Jember. The animals were kept in separate cages, one cage containing one rat, using a plastic box (45×30×15 cm3) with woven wire as a cover. The floor mat was covered with wood shavings and an underpad was placed to absorb urine and maintain moisture. The light sources were a 300-lux electric lamp with a distance of 1 m from the floor. The cage was cleaned every 3 days with soap and running water. The room comfort was provided by air conditioning to maintain a room temperature of 22°C±2°C and humidity of 50%–70%. An exhauster fan was used to remove the smell of ammonia. The environment was a quiet room with 12 hours of light and dark cycle. The feed was 30–35 g of pellets (10% of body weight) and 30–35 mL of mineral water (10% of body weight).
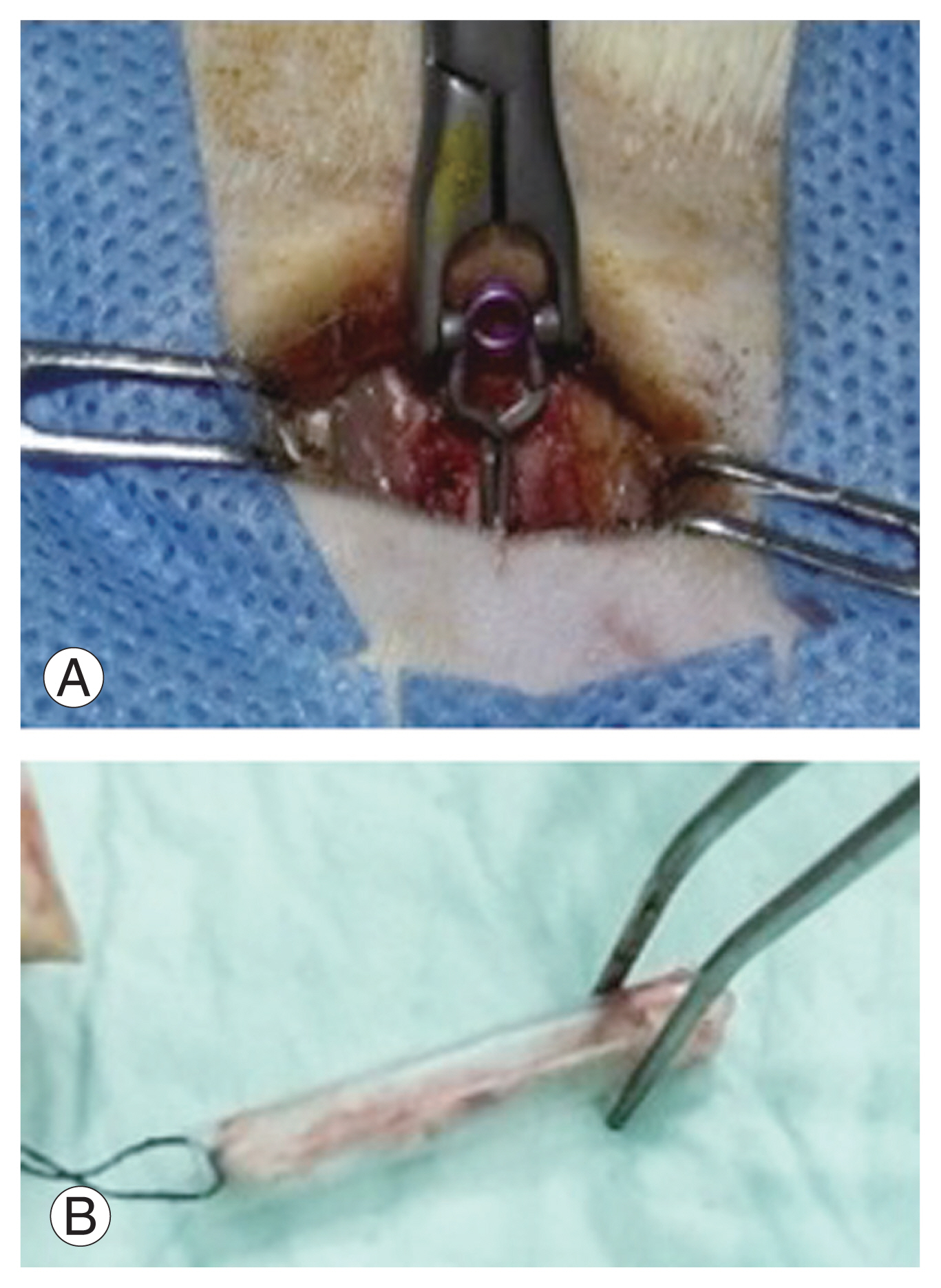
Contusion-compression of the spinal cord was conducted by a commercially available spinal cord impactor, aneurysm Yasargil clip, with a length of 7 mm and load of 65 g (equivalent to 150 kdyn). The animals were anesthetized using ketamine (75 mg/kg) and acepromazine (3 mg/kg) intraperitoneally [15]. Moreover, they were placed on a fixation board in the prone position, and approximately 3 cm of their back hair was shaved. The operating area was disinfected with 10% betadine and 75% alcohol. The surgical level was marked by tracing the Th12 rib to the Th12 spinous process to create a 2 cm skin incision. T10–T11 laminectomy exposed the T10 spinal cord. The tip of the titanium aneurysm Yasargil clip was placed at a distance of 1 mm from the spinal cord anteriorly and posteriorly. The spinal cord was impacted suddenly for 60 seconds by the retraction of the tip of the aneurysm Yasargil clip using an applicator. It produced a SCI contusion-compression model with the dura appearing cloudy white and impacted flat. The operating field was cleaned using saline, and the muscle and skin were sutured together in layers.
At 3 days post-injury, the rat SCI subacute contusion-compression models in the secretome and non-secretome groups were paraplegic (0). The SCI+secretome group was given an intrathecal injection of 30 μL HNSC secretomes under general anesthesia, centered at the site of injury, 1.5–2 mm deep from the dura to the subarachnoid space at a tilt angle of 30°–40° using a 50 μL Hamilton syringe. The correct position of the needle tip in the intrathecal space between the dura matter and the spinal cord should be determined while avoiding the risk of spinal cord trauma by the needle tip. Once the needle penetrates the dura matter, the cerebrospinal fluid is aspirated, becomes clear, is mixed with HNSC secretome, and is finally injected into the intrathecal space. The animals were administered with physiological saline subcutaneously (s.c.), tolfenamic acid (4 mg/kg s.c.), and enrofloxacin (10 mg/kg s.c.) and exposed to a 5-watt heating lamp. Manual bladder drainage was performed twice daily until micturition was normal.
5. Functional assessment
The assessments of the locomotor expression and BBB open-field tests were performed on days 1, 3, 7, 14, 21, 28, and 35 after injury [16]. The tail, body, legs, trunk stability, limb movement, and toe clearance were examined based on the BBB scores to measure the locomotor abilities. The results indicated the numbers ranging from 0 to 21. A score of 0 indicates no movement, while a score of 21 indicates normal movement without locomotor disorders.
6. Preparation of tissue for immunohistochemical assessment
The termination of animals was performed on day 35 using inhalation anesthetics. Ether was dropped on a cotton swab and placed with the animals in a closed glass jar. The activity of the animals was then observed. The animals began to fall asleep, as indicated by dimming eye activity, and death was indicated by no breathing or heart rate and pale white eyes. The sterilization of the operating area was performed using 70% alcohol and 10% betadine and sterile cloths. The animals were prepared in the prone position. Th8–Th12 were identified by palpating the ribs and then traced to the transverse spine process. A transverse incision was made on the left side of the ribs reaching the Th8–Th12. Moreover, a 5 cm SCI was separated from the vertebral column and marked at the cranial end. The SCI material obtained from the debridement procedure was put in a pot and fixed in 10% buffer formalin to be sent for immunohistochemical examination at the anatomical pathology laboratory (Fig. 1).
7. Immunohistochemical assessment
Immunohistochemical MMP9, TGF-β, and BDNF were evaluated via quantitative measurement. Five spinal cord tissue specimens were harvested from the animals in each group. We observed an average value of 10 fields of view, every field of view having 625 μ2 with 400× magnification. The process of immunohistochemical evaluation involves 12 steps: (1) fixation, (2) embedding and sectioning, (3) deparaffinization and hydration, (4) antigen retrieval, (5) blocking, (6) addition of primary antibodies, (7) incubation, (8) washing with Tris-buffered saline with Tween 20, (9) second washing, (10) addition of substrates, (11) third washing, and (12) counter straining.
8. Statistical methods
The data in this research are reported as the mean±standard deviation of the mean. The IBM SPSS software ver. 25.0 (IBM Corp., Armonk, NY, USA) was used to analyze the differences between the groups by analysis of variance followed by Tukey’s post hoc test. A p-value of <0.05 was considered statistically significant.
Results
1. The Basso–Beattie–Bresnahan locomotor score
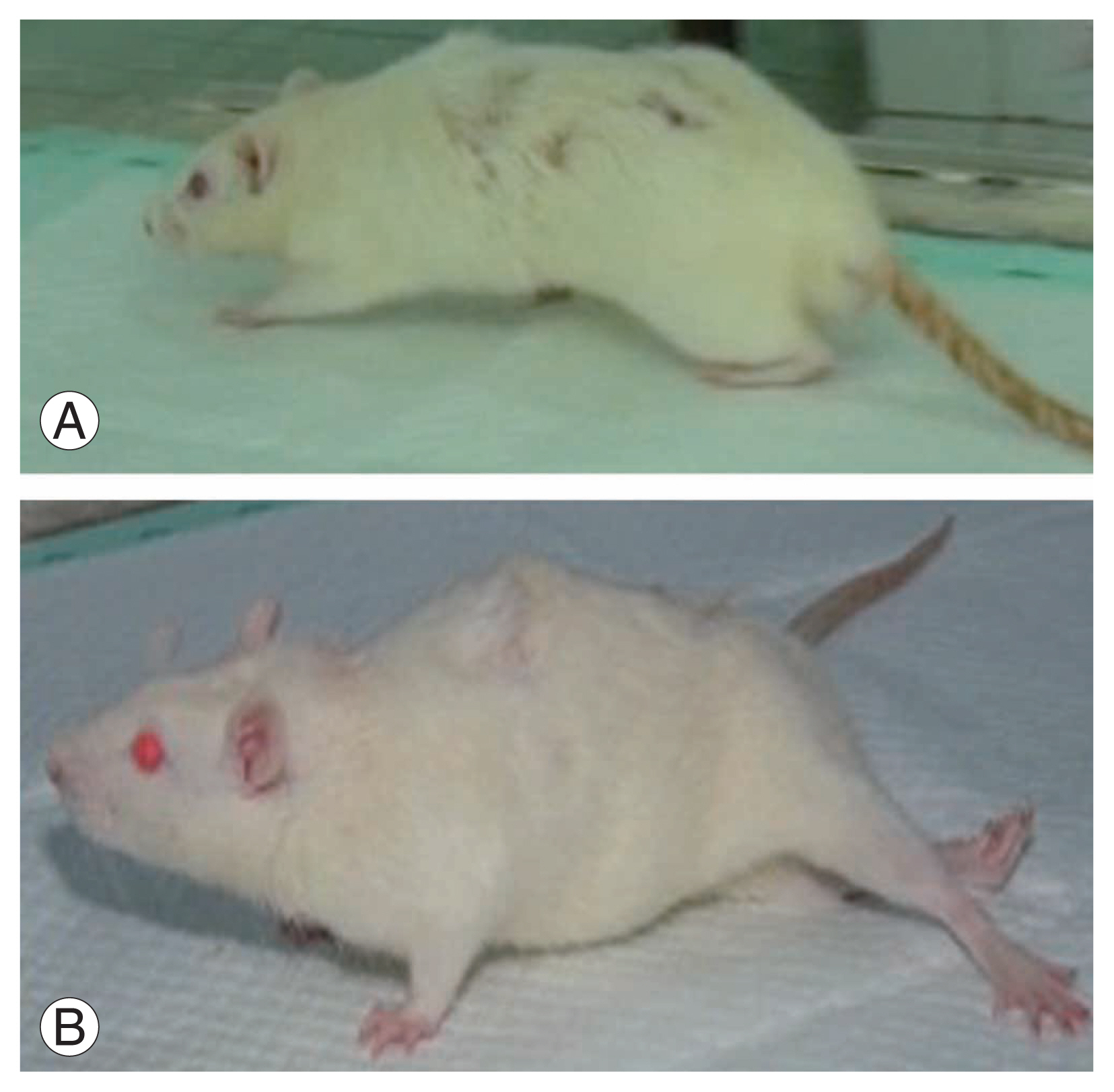
The rats were examined over a period of 5 weeks to assess the recovery of their motor function. Locomotor recovery started on day 7 and continued until day 35. The mean BBB score in the SCI+non-secretome group was 1.8, while that in the SCI+secretome group was 20.2, with a mean difference in the BBB scores of 18.4 (BBB score range, 0–21). The results of the independent t-test showed that the non-secretome and secretome groups had a significant difference of 0.000 (p<0.05) (Figs. 2, 3).
2. Immunohistochemical interleukin-10
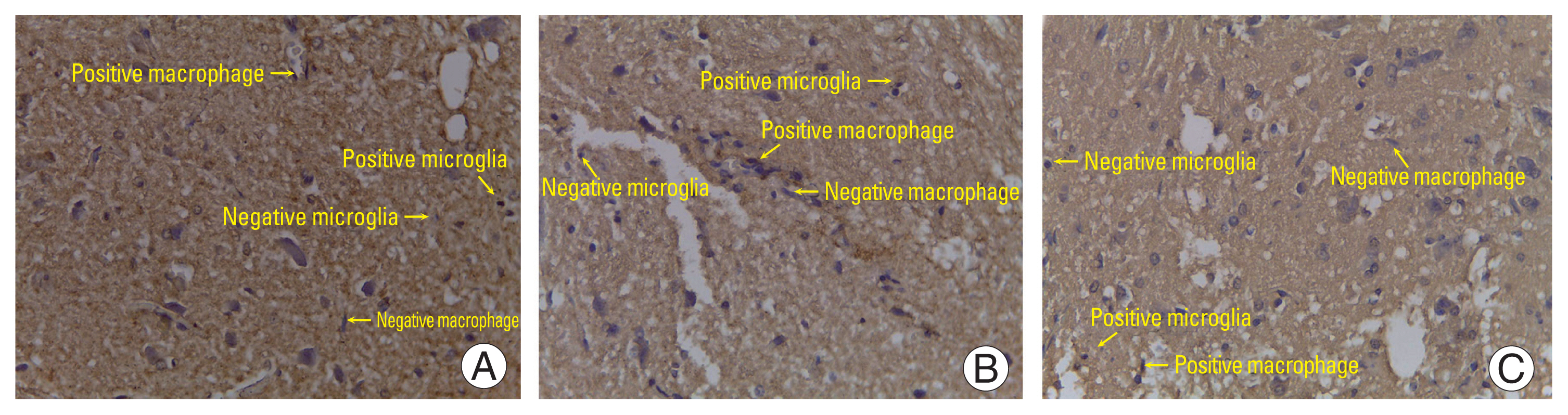
The number of cells expressing IL-10 in the normal group was smaller than that in the SCI+secretome group (p<0.05). The number of cells expressing IL-10 in the normal group was not significantly different from that in the SCI+non-secretome group (p>0.05). The number expressing IL-10 in the SCI+secretome group was significantly different from that in the SCI+non-secretome group (p<0.05), while the number of cells expressing IL-10 in the SCI+secretome group was greater than that in the SCI+non-secretome group (Table 1, Fig. 4).
3. Immunohistochemical transforming growth factor-β
The number of cells expressing TGF-β in the normal group was significantly different from that in the SCI+secretome group (p<0.05). On the other hand, the number of cells expressing TGF-β in the normal group was not significantly different from that in the SCI+non-secretome group (p>0.05), but the number of cells expressing TGF-β in the SCI+non-secretome group was greater than that in the normal group. The number of cells expressing TGF-β in the SCI+secretome group was significantly different from that in the SCI+non-secretome group (p<0.05) (Table 2, Fig. 5).
4. Immunohistochemical matrix metalloproteinase 9
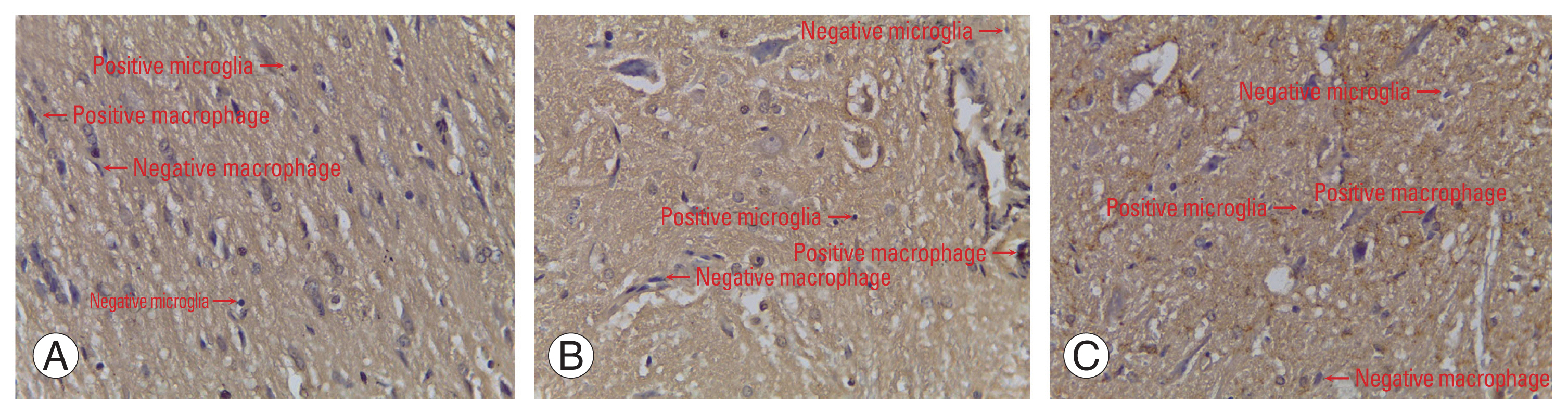
The number of cells expressing MMP9 in the normal group was not significantly different from that in the SCI+secretome group (p>0.05), while the number of cells expressing MMP9 in the normal group was significantly different from that in the SCI+non-secretome group (p<0.05); the number of cells expressing MMP9 in the normal group was smaller than that in the SCI+non-secretome group. The number of cells expressing MMP9 in the SCI+secretome group was significantly different from that in the SCI+non-secretome group (p<0.05); the number of cells expressing MMP9 in the SCI+secretome group was smaller than that in the SCI+non-secretome group (Table 3).
Discussion
Permanent neurodeficit and glial scarring can be caused by trauma to the spinal cord [1,2]. After SCI, the major causes of the regenerative failure of injured axons include the formation of a glial scar and inflammation [2,3]. Stem cell secretomes are non-cell metabolites containing cytokines, chemokines, growth factors, extracellular vesicles, or exosomes [10]. HNSC secretomes can have the size of up to 1,000 nm, derive from cell secretion/shedding, are collected from the cell culture media, and comprise a combination of other components, whereas HNSC exosomes can have the size of up to 200 nm; derive from multivesicular body pathways; are collected by filtration, ultracentrifugation, precipitation, and immunoaffinity; and comprise receptors, transcription factors, enzymes, proteins, lipids, and nucleic acids (DNA, mRNA, and miRNA) [10]. Stem cell secretomes are potentially neuroinflammatory, anti-apoptotic, angiogenic, and neurogenic [7,10]. The administration of secretomes in the brain could induce macrophages to secrete IL-10 and TGF-β and decrease MMP9 [7]. The effect of HNSC secretome administration on spinal cord repair remains unclear. Accordingly, this study aims to investigate the effects of treatment with HNSC secretomes on subacute SCI post-laminectomy by analyzing IL-10, MMP9, TGF-β, and BBB score locomotors as expressions of neurological recovery.
We showed that the treatment with HNSC secretomes for the SCI of rats’ post-laminectomy significantly improved locomotor recovery that started on day 7 and continued until day 35 based on the BBB scores (p=0.000, mean=18.4) compared with the SCI+non-secretome group. The HNSC secretome decreased MMP9 (p=0.015) but increased IL-10 (p=0.045) and TGF-β (p=0.01) compared with the SCI+non-secretome group.
IL-10, an anti-inflammatory cytokine, reduces the synthesis of MMP9, induces the polarization of the M2 macrophage phenotype, reduces inflammatory response, and suppresses inflammatory cells [17–19]. Moreover, it can inhibit the early effects of MMP9 in terms of the degradation of the BSCB matrix basal lamina [2,17]. The inhibition of glial scar formation is caused by decreased astrocyte migration, dialog between infiltrating macrophages, and astrocyte-mediated extracellular matrix (ECM) deposition [20]. Previous studies have shown that systemic IL-10 injection results in significant neuroprotective and improved functional recovery after traumatic SCI [18]. IL-10 also provides trophic support to neurons through the downregulation of the pro-apoptotic factors caspase 3, cytochrome c, and Bax and the upregulation of B-cell lymphoma (Bcl) anti-apoptotic factor 2 (Bcl-2) and X-linked Bcl-2 and Bcl-xl [18,21]. These positive effects include the reduction in lesion size and motor recovery improvement [18].
TGF-β, an anti-inflammatory cytokine, induces regulatory T-cell differentiation and promotes macrophage polarization toward the M2 phenotype [21]. It prevents M1 from synthesizing with proinflammatory TNF-α, IL-1β, IL-6, inducible NO synthase, reactive oxygen species, glutamates, and proteases [22]. Moreover, TGF-β induces fibroblasts to secrete collagen, fibronectin, and proteoglycans that results in new ECM formation [23]. TGF-β overcomes the matrix degradation, which is caused by the effects of MMP9 [24]. TGF-β is involved in nerve repair and regeneration; combating nerve damage; stimulating cellular survival, growth, proliferation, and differentiation; and regulating the behavior of neurons and glial cells [25]. This research showed that the HNSC secretome increases TGF-β in the rat SCI subacute contusion-compression model.
MMP9 are endopeptidases that contribute to SCI repair by degrading the ECM molecules and depleting MMP9 [26]. MMP9 are inhibited by the tissue inhibitors of metalloproteinases (TIMP), while TIMP are inhibited by TGF-β [25]. MMP9 is responsible for the early degradation of the basal lamina components, which leads to BSCB disruption and initiates leukocyte infiltration, progressive neuroinflammation response, and demyelination [27]. This system modulates macrophage invasion and myelin destruction, which has an important role in neuropathic pain [2]. Moreover, MMP9 regulates neuropathic pain through glial scar formation [6]. This research showed that HNSC secretome decreases MMP9 in the rat SCI subacute contusion-compression model.
This study showed that HNSC secretome can repair subacute SCI post-laminectomy. It substantially improved locomotor recovery and increased the levels of IL-10 and TGF-β but decreased MMP9. IL-10 decreased the matrix degradation caused by MMP9 inhibition, while TGF-β increased the locomotor function and new ECM formation via fibroblast-induced collagen synthesis, reduced glial scar formation, and axon sprouting.
The limitations of this study include a small sample size and the analysis of neuroinflammatory and ECM factors. Future studies could increase the number of experimental animals and investigate the regeneration mechanism and observe the biomarker variables (i.e., neuroinflammation, neuroapoptosis, neuroangiogenesis, neurogenesis), histopathological variables, and expression variables in relation to locomotor function.
Conclusions
The effect of the treatment with HNSC secretomes on subacute SCI post-laminectomy in R. norvegicus rats substantially improved locomotor recovery and increased the levels of IL-10 and TGF-β but decreased MMP9. The factors of the HNSC secretomes can mitigate the pathophysiological processes of secondary damage after SCI and improve locomotor functional outcomes in R. norvegicus.