Introduction
During posterior spine surgery, spine surgeons always encounter yellowish epidural fat (EF) that covers the dura mater and the nerve root after ligamentum flavum (LF) resection. However, the role and function of EF remains unclear. Spine surgeons have limited knowledge regarding EF; therefore, no effort is made to preserve it during surgical procedures, resulting in its loss and exposure of the dura mater and the nerve roots [1].
What is the role of EF, and why is it located between the LF and the dura mater (spinal cord) or the nerve root? Few anatomical and histologic studies have concluded the following: (1) EF is also an adipose tissue; however, its histologic morphology and shape differ from those of subcutaneous fat [2-5]; and (2) the EF may enable smooth sliding of the nervous structures, such as the dura mater and the nerve roots from the LF and other stable structures [1,5,6]. However, to our knowledge, no studies have investigated EF in detail, especially focusing on the mesenchymal stem cells (MSC) in the EF tissue.
The authors hypothesized that the EF may have critical functions for the spinal cord, nerves, dura mater, and surrounding structures. In addition, if EF is in fact an adipose tissue, MSC with specific regenerative and anti-inflammatory functions for the surrounding structures may be present [7-10]. According to this hypothesis, we have conducted several experimental studies on EF. The current study, the first and fundamental report of these experimental and clinical studies, focuses on ŌĆśwhether EF tissue contains MSC.ŌĆÖ
Materials and Methods
1. Epidural fat-derived stem cell isolation and culture
This study was approved by the Institutional Review Board and Ethical Committee of the Yeungnam University Medical Center (YUMC IRB no., 2017-07-032). The EF was obtained from consenting patients (n=3) during posterior decompression surgery of the lumbar spine performed at our center. The specimens were placed in sterile tubes, stored at 4┬░C, and brought to the Laboratory Animal Center of the Daegu-Gyeongbuk Medical Innovation Foundation to be processed within 12 hours.
The primary cells were isolated and cultured as previously described with some modifications [11]. Briefly, the collected fat tissues were washed with phosphate-buffered saline (PBS; Gibco Invitrogen, Carlsbad CA, USA) 3ŌĆō4 times to remove blood and debris. The washed tissues were minced using scissors and surgical blades. The tissues were incubated with collagenase type I (2 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) for 1ŌĆō2 hours. The digested samples were sieved with a cell strainer (pore size=70 ╬╝m, Falcon; BD Biosciences, Coning, NY, USA) and then washed with PBS and centrifuged at 350 ├Śg for 10 minutes for cell isolation. The cell pellet was suspended in low glucose DulbeccoŌĆÖs modified EagleŌĆÖs medium (Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum (Invitrogen). The cells were cultured in a 5% CO2 incubator with culture medium changes thrice a week.
2. Cumulative population doubling level analysis
The proliferation ability of the cultured cells was measured, as previously described, with some modifications [12,13]. The growth and proliferation ability of the cultured cells were estimated via the cumulative population doubling level (CPDL) using the formula CPDL=ln (Nf/Ni) ln2, where Ni is the initial number of cells seeded, Nf is the final number of harvested cells, and ln is the natural log. The cells (3├Ś105) were seeded in triplicate using 6-well culture dishes and subcultured for 3 days. Thereafter, the cells were counted and reseeded. To estimate the CPDL, the population doubling of each passage was measured and added to the previous cell population double levels.
3. Reverse transcriptionŌĆōpolymerase chain reaction
Total RNA was extracted from the cultured cells using an RNeasy mini kit and RNase-Free DNase set (QIAGEN, Hilden, Germany) as per the manufacturerŌĆÖs protocol. RNA concentrations were measured by the ratio of absorbance at 260 nm and 280 nm using a spectrophotometer. Complementary DNA (cDNA) was prepared using 1 mg of total RNA for reverse transcription (RT) using Super-Script III reverse transcriptase (Invitrogen, Groningen, Netherlands). The cDNA was amplified with polymerase chain reaction (PCR) with Takara Ex Taq (Takara Bio, Kusatsu, Japan). The PCR primers are shown in Table 1. The PCR products were separated on a 1.5% agarose gel and visualized with ethidium bromide. Quantitative RT-PCR was performed by mixing the cDNA with primers and SYBR Green I Master Mix (Roche Diagnostics, Mannheim, Germany). Quantitative RT-PCR was performed using a LightCycler 480 system with supplied software (Roche Diagnostics), as per the manufacturerŌĆÖs instructions. The RNA expression levels were compared after normalization to endogenous glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
4. Fluorescence-activated cell sorting analysis
To determine the expression of cell surface markers, the cultured cells were stained and analyzed using flow cytometry. Cell staining was performed as per the manufacturerŌĆÖs cell surface immunofluorescence staining protocol (Biolegend, San Diego, CA, USA). Briefly, the cells were washed twice with PBS and detached using 0.25% trypsin/ethylenediaminetetraacetic acid. The detached cells were collected, washed with PBS, and divided into groups for antibody staining. Each group contained about 1├Ś105 cells for staining. The antibodies used to detect the cell surface markers included CD105, CD90, CD73, CD45, CD34, and CD14 (BioLegend; all antibodies for anti-human). All the antibodies were conjugated with fluorescein dye. The staining was conducted for 30 minutes at 4┬░C with antibody incubation. After the incubation, the cells were washed twice with PBS. Fluorescence-activated cell sorting (FACS) analysis was performed with a FACS Calibur cytometer (BD Biosciences, San Jose, CA, USA).
5. Osteogenesis
To confirm the osteogenic differentiation ability of human EF-MSC, the cells were treated with an osteogenic differentiation medium (StemPro Osteogenesis Differentiation kit, Gibco Invitrogen). For osteogenic differentiation, the cells were cultured in 6-well culture plates. At 70%ŌĆō80% cellular confluence, the culture medium was changed to an osteogenic differentiation medium. The differentiation medium was changed twice weekly for 3 weeks. The differentiated cells were stained with von Kossa and alizarin red S staining to confirm calcium deposition. For von Kossa staining, the cells were stained with 5% silver nitrate for 1 hour and exposed to ultraviolet light. Following incubation, the differentiated cells were treated with 5% sodium thiosulfate for 3 minutes and counterstained with nuclear red stain for 3 minutes. For alizarin red S staining, the differentiated cells were fixed with 70% ethanol for 1 hour at 4┬░C. After fixation, the cells were washed with distilled water and stained with alizarin red S (Sigma-Aldrich) for 10 minutes at room temperature. Thereafter, the cells were washed 3ŌĆō4 times with distilled water. Alizarin red SŌĆōstained cells were solubilized in cetylpyridinium chloride (Sigma-Aldrich) for 1 hour. A spectrophotometer was used to measure the absorbance of the solubilized cells at 570 nm.
6. Adipogenesis
To confirm the adipogenic differentiation ability of human EF-MSC, the cells were treated with adipogenic differentiation medium (StemPro Adipogenesis Differentiation kit, Gibco Invitrogen). When the cells reached 70%ŌĆō80% confluence, the adipogenic differentiation medium was used for 3 weeks. After adipogenic differentiation, oil red O staining was performed to detect the lipid droplets. The cells were fixed with 10% formalin for 1 hour and washed with 60% isopropanol. After fixation, the cells were stained with oil red O for 10 minutes. The oil red O-stained cells were solubilized in 100% isopropanol, and the absorbance of the solubilized solution was measured at 500 nm using a spectrophotometer.
7. Chondrogenesis
To detect the chondrogenesis differentiation ability of human EF-MSC, the cells were treated with chondrogenic differentiation medium (StemPro Chondrogenesis Differentiation kit, Gibco Invitrogen). The cells (2├Ś105) were seeded in a 15-mL polypropylene tube and spun down in a centrifuge. The cells were treated with the chondrogenic differentiation medium (1 mL) for 3 weeks. The differentiation medium was changed 3 times every week. After differentiation, the cells were confirmed the pellet formation. The pellet was embedded in a paraffin block, sectioned, and stained with toluidine blue to confirm the chondrogenic differentiation ability.
Results
1. Stem cell properties of epidural fat-derived mesenchymal stem cells
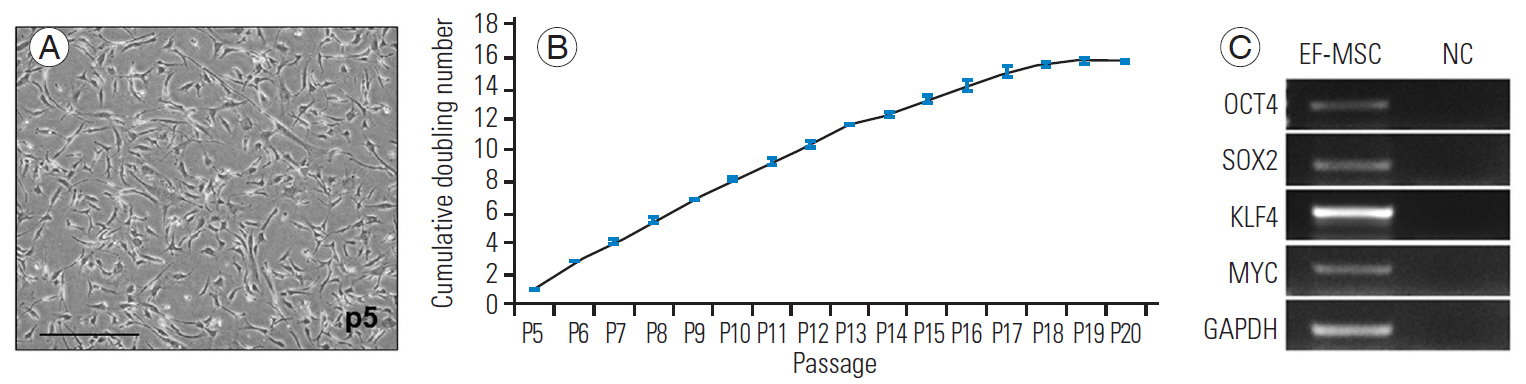
After 3 weeks of culturing of MSC isolated from human EF tissues (EF-MSC), plastic-adherent cells with a unique morphology were obtained. The EF-MSC exhibited neuronal or fibroblast-like bipolar morphologies until senescence at p18 (Fig. 1A). Throughout the culturing, they underwent a mean 15.8 doublings and produced 6.9├Ś1016 cells (Fig. 1B).
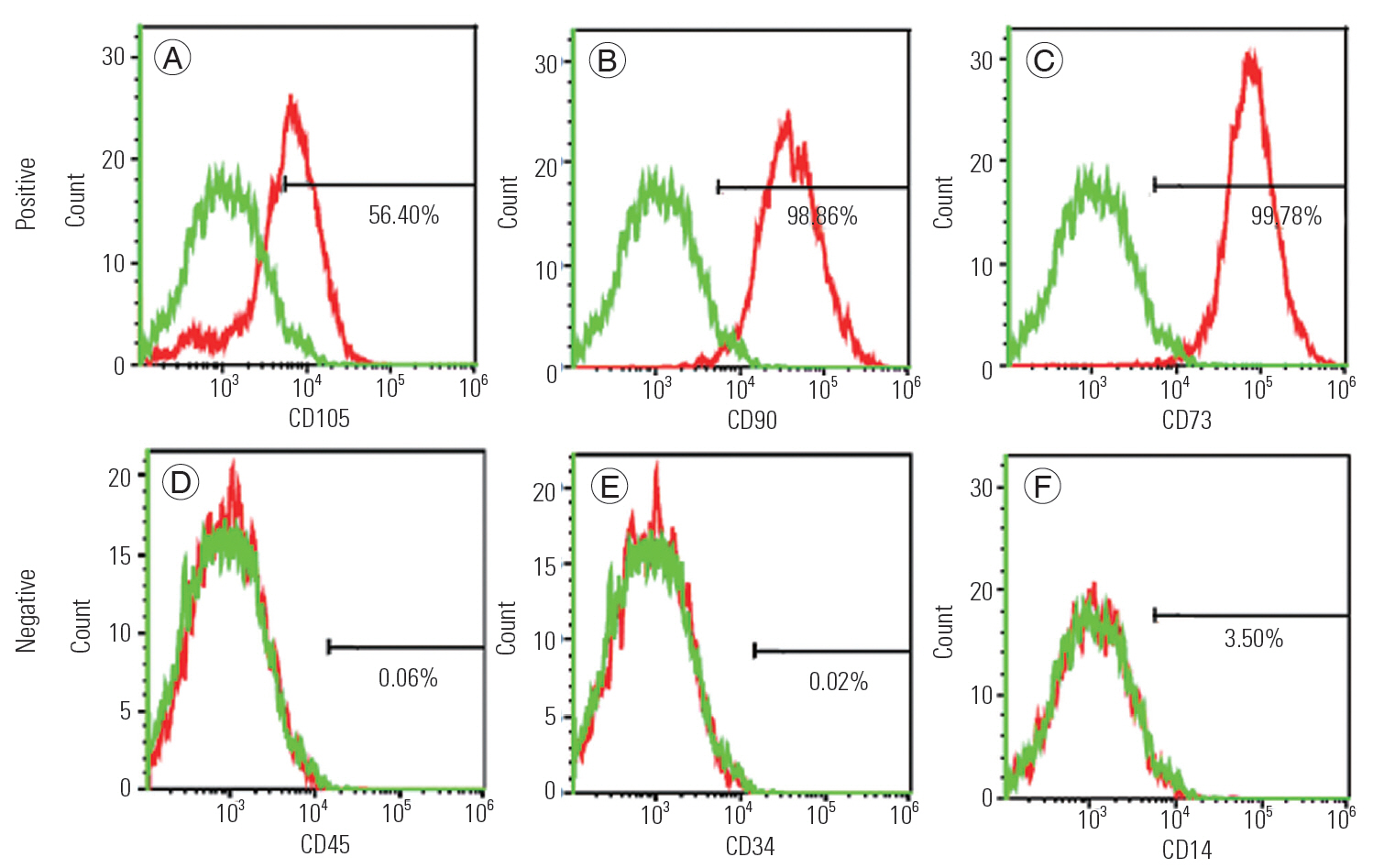
In the RT-PCR analysis, EF-MSC at p4 expressed stem cell-specific genes, including OCT4, SOX2, KLF4, MYC, and APDH; however, no expression was observed in the control group (Fig. 1C). The FACS analysis showed that EF-MSC at p4 expressed stem cell-specific surface markers of CD105, CD90, and CD73 but not those of CD45, CD34, and CD14 (Fig. 2).
2. Differentiation potentials of epidural fat-derived mesenchymal stem cells
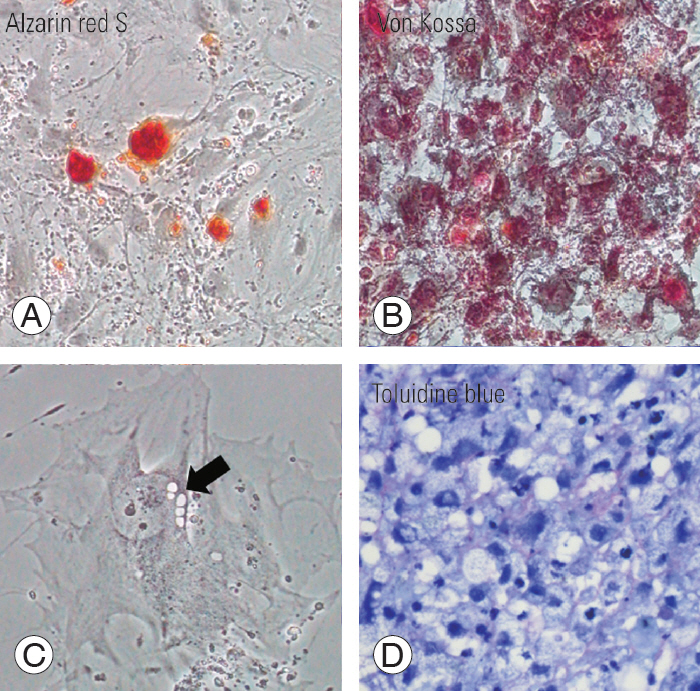
EF-MSC cultured in appropriate differentiation media showed osteogenic (with alizarin red S stain and von Kossa stain), adipogenic (with oil red O stain), and chondrogenic (with toluidine blue stain) differentiation potentials (Fig. 3).
Discussion
Currently, spine surgeons remain unaware regarding what EF tissue is and why it is located in the space between the LF and the neural structures. However, EF tissues are eliminated completely after surgical procedures, and spine surgeons have suggested that full exposure of the dura mater and nerve root with EF tissue removal indicate full decompression and surgical success. However, several studies have documented some problems after spine surgery, such as post-laminectomy syndrome, caused to the lack of knowledge among surgeons about the role of EF tissue [1,2,5,6]. We hypothesized that the EF tissue may also play a significant role in the neural and other nearby structures. Thus, we conducted several experimental and clinical studies on EF tissue. The current fundamental and basic study focused on whether EF tissues contain EF-MSC.
To certify whether the cells derived from certain tissues are MSC, several steps need to be completed. The current study revealed that the cells had a fibroblast- or neuron-like morphology typically observed in MSC that remained until senescence at p18. MSC-specific genes such as OCT4, SOX2, KLF4, MYC, and GAPDH were expressed in the RT-PCR analysis, and MSC-specific surface markers, such as CD105, CD90, and CD73 were exhibited in the FACS analysis. Finally, and most importantly, the cellsŌĆÖ adipogenic, chondrogenic, and osteogenic differentiation properties were verified (Fig. 3). Based on the outcomes, we concluded that EF tissues possess MSC (EF-MSC), and the EF-MSC may have specific regenerative properties for the nervous or other nearby structures and anti-inflammatory properties for the epidural space or the surrounding structures.
Thus far, few studies have evaluated the histologic features of EF tissues or its impact on epidural adhesion or scarring [2,4,5]; however, no studies have investigated EF-derived MSC and their specific functions. The MSC derived from adipose tissues have the following two specific functions; regenerative effect and an anti-inflammatory effect that are more effective for nearby structures [7-10,14]. Thus, EF-MSC may perform some of the following functions. First, the EF-MSC may impact the regeneration of nearby neural structures, such as the spinal cord. Spinal cord injuries are occasionally induced by trauma or disease; however, no definite treatment method has been established for them. Many experimental studies using MSC have been conducted [15-17]; however, they are mostly induced from subcutaneous fat tissue owing to the ease in harvesting and availability, or other originated stem cells. Second, EF-MSC may also affect peridural adhesion or scarring by MSC-induced anti-inflammatory properties [10,16,18,19]. Postsurgical epidural scarring or adhesions are considered significant issues for clinical impairments, such as the post-laminectomy syndrome [1,6] that may be caused by the removal of the EF tissue during spine surgery. Considering the anti-inflammatory effects of MSC, the preservation of EF tissue may reduce specific epidural adhesion-related conditions. In fact, the authors have performed several related experimental and clinical studies, and we shall present the findings in the near future.
The current study had certain limitations. First, we described the stem cell features of EF-MSC without comparing it to other MSC, such as subcutaneous tissue-derived MSC. Most MSC studies conducted thus far have used subcutaneous tissue-derived MSC in fat tissues; therefore, the comparison between the EF-MSC and those derived from subcutaneous tissue may be instructive. We are aware of this limitation; therefore, we intend to conduct a comparative study of the two MSC in the future. Second, the unique features of EF-MSC should be investigated experimentally in future studies. Moreover, as described in the ŌĆśDiscussionŌĆÖ section, the EF tissue may have a critical influence on the patientsŌĆÖ clinical outcomes. The current study was fundamental for further experimental and clinical studies regarding EF-MSC; therefore, additional studies are important for defining its significance. Despite these limitations, the study has significant strengths. This was the first study to identify EF-MSC that may be an important stem cell source for specific pathologies and may have a critical influence on spine conditions. Thus, we believe that this study can be a cornerstone for further studies on EF-MSC.
Conclusions
To our knowledge, this is the first study to investigate EF-MSC. Based on the cell culture, FACS analysis, RT-PCR analysis, and differentiation potential (adipogenesis, chondrogenesis, and osteogenesis) outcomes of this study, all the features of the cells corresponded to MSC, and we defined EF-MSC in the EF tissue. Further experimental and clinical studies with EF-MSC are warranted to identify the significance of our findings.